Zusatzinformationen
Bezeichnung der REs | Häufigkeit der Schnitte | Das RE-Problem | star activity | Isoschizomere | Typen des Restriktionsverdaus | rekombinante Herstellung von REs
Es sind drei Klassen von Restriktionsendonukleasen bekannt. Die Enzyme der Klasse II sind die am häufigsten eingesetzten. Sie haben folgende Eigenschaften, die sie in der Anwendung so wertvoll mach
- sie besitzen eine Erkennungssequenz, innerhalb der auch geschnitten wird
- die Erkennungssequenzen bestehen aus 4, 6 oder 8 Nukleotiden
- die Erkennungssequenzen sind spiegelbildlich ("palindromisch"), d.h. sie ergeben von vorne und hinten gelesen die gleiche Sequenz
- sie benötigen außer Magnesium-Ionen keine weiteren Effektoren
| Typ I | Typ II | Typ III | |
| Funktion | Endonuklease + Methylase | Endonuklease | Endonuklease + Methylase |
| Erkennungssequenz | 2-teilig, asymmetrisch | 4-8 Basen, palindromisch | 5-7 Basen, asymmetrisch |
| Spaltstelle | unspez., bis 1000 Bp vor der Erkennungsstelle | innerhalb der Erkennungsstelle | 5-20 Basen vor der Erkennungsstelle |
| ATP-Bedarf | ja | nein | ja |
Beispiele für Restriktionsendonukleasen des Typ II:

Abbildung 1: Beispiele für RE-Schnitte in Erkennungssequenzen.
Bezeichnung der Restriktionsendonukleasen
Da die Restriktionsendonukleasen aus Bakterien stammen, werden sie in der Regel nach ihrem Keim benannt. Am Beispiel von Eco RI: E=Gattung (Escherichia), co (coli) = Art, R = Stamm, I erste RE
Häufigkeit der Schnitte von Restriktionsendonukleasen
Es kann erwartet werden, dass mit zunehmender Länge der Erkennungssequenz diese Nukleotidfolge weniger häufig in einer DNA vorkommt und somit die Häufigkeit der Schnitte abnimmt. Die Statistik hilft bei der Abschätzung dieser Schnitthäufigkeit.
Bei 50% GC-Gehalt gilt: 1 Schnitt pro 4n Nukleotide, wobei
4 für die Anzahl der Nukleotidbasen A,T,G und C in der DNA und
n für die Anzahl der Nukleotide in der Erkennungssequenz steht.
Für n=4 (bei Hae III, Alu I) ergibt sich also nach 44=256 Nukleotiden ein Schnitt.
Für n=6 (Eco RI und viele andere RE) ergibt sich nach 46=4096 Nukleotiden ein Schnitt; für n=8 ("rare cutter" wie Not I) haben alle 48 = 65536 Nukleotide einen Schnitt.
Das RE-Problem
Wenn also Restriktionsenzyme DNA zerschneiden, warum wird dann nicht das Chromosom der Produzenten zerschnitten? Oder: wie kann das Bakterium zwischen der Fremd-DNA aus dem Phagen und der eigenen DNA unterscheiden?
Die eigene DNA wird in den Erkennungssequenzen des eigenen Restriktionsenzyms an Cytosin oder Adenin methyliert. Diese methylierenden Enzyme kodieren in der Regel zusammen mit dem Restriktionsenzym. Sie werden in das dam (methyliert Adenin) und das dcm (methyliert Cytosin) eingeteilt. Damit kann das Restriktionsenzym diese modifizierte Schnittstelle nicht mehr erkennen. Es wird also von einem Restriktions-Modifikationssystem gesprochen.
Die praktische Bedeutung dieses Systems ist leicht bei der rekombinanten Herstellung der Restriktionsendonukleasen zu erkennen. Würde die Restriktionsendonuklease Hae III in E. coli hergestellt, so müsste nicht nur das Gen HaeIII, sondern auch das Gen für die zugehörige Methylase (M.HaeIII) mit einkloniert werden. Siehe dazu eine Originalpublikation (pdf-Datei, 270 KB)zum Eco RI.
Star activity
Restriktionsendonukleasen sind Phospodiesterasen, die in ihrer Aktivität beeinflußbar sind. Niedrige Salzkonzentrationen, hoher pH (>8), Glycerin und organische Lösungsmittel (Ethanol, Dimethylsulfoxid, etc.) können das Enzym veranlassen, gar nicht oder an anderen Stellen die DNA zu hydrolysieren. Für viele REs sind Bedingungen wie die folgenden geeignet: 30-50 mM Tris-Acetat pH 7.5, 10 mM Mg-Acetat, 50-100 mM K-Acetat und 0,5 mM DTE. DTE oder DTT sind Reduktionsmittel und 3-5 U der RE. Ein solcher Ansatz bildet das Cytoplasma einer Bakterienzelle nach.
Isoschizomere/Neoschizomere
Um methyliert Schnittstellen zu spalten, werden isoschizomere REs verwendet. Solche Isoschizomere sind weniger empfindlich gegenüber der Methylierung. Sie schneiden an der gleichen Stelle, wie die durch Basen-Methylierung gehemmte RE. Beispiel: Bst I ist das Isoschizomer von Bam HI. Beide haben die Erkennungssequenz (G/GATCC); sie schneiden an der angegeben Stelle. Neoschizomere haben die gleiche Erkennungssequenz, schneiden aber an anderer Stelle innerhalb dieser Sequenz. Informationen zu solchen Isoschizomeren entnehmen Sie bitte der REBase-Datenbank oder den Katalogen der Enzymlieferanten.
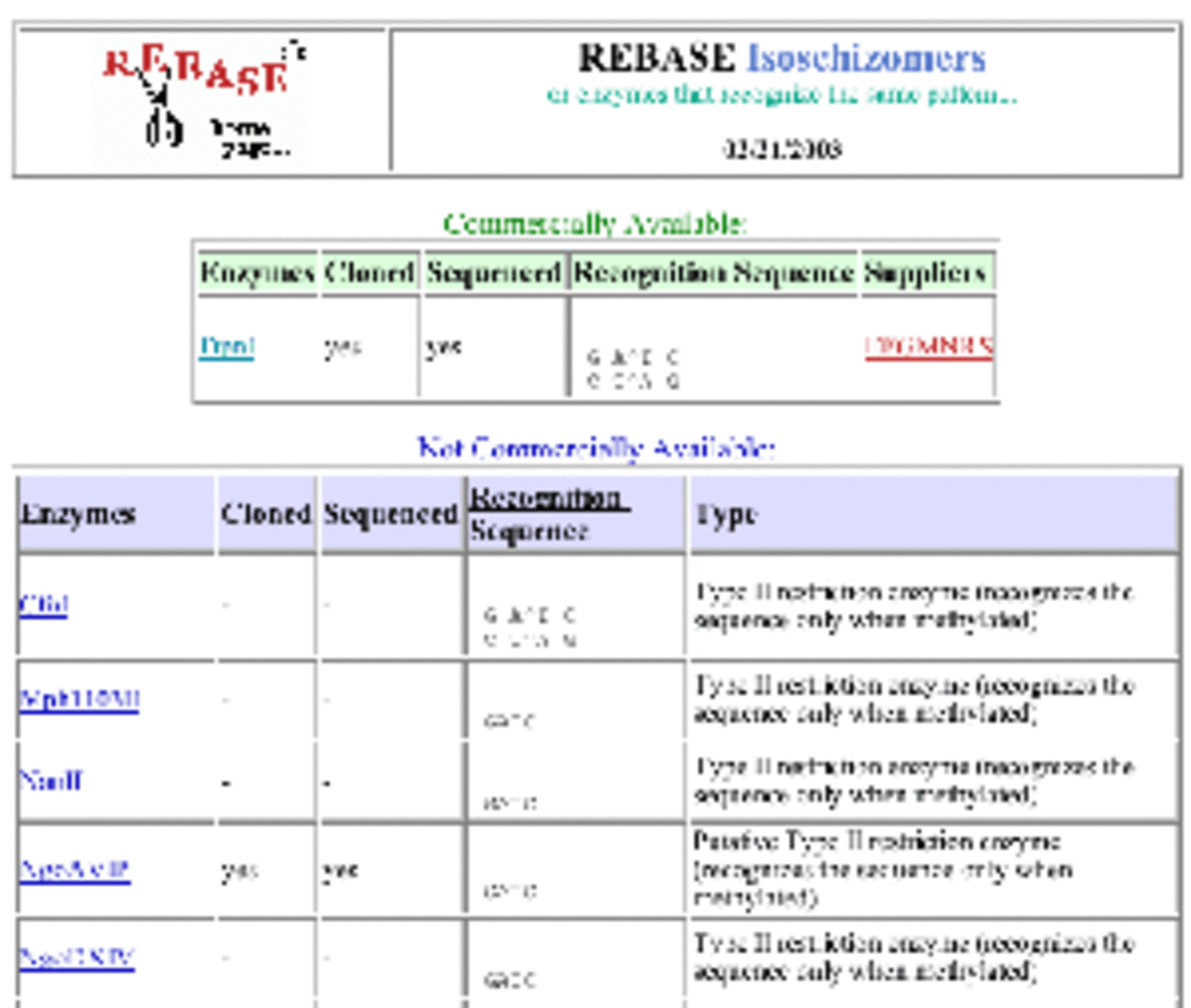
Abbildung 2: Beispiel aus der REBase-Datenbank zur Suche nach Isoschizomeren von Dpn I.
Typen des Restriktionsverdaus
Man unterscheidet den vollständigen Verdau vom Partialverdau und vom Mehrfachverdau. Der vollständige Verdau, also die Hydrolyse aller vorhandenen Schnittstellen einer RE, ist bei Plasmidanalysen und bei Restriktionslängen Polymorphismen erwünscht. Ein Partialverdau durch REs mit 4-er Erkennungsstelle wie Sau 3A (/GATC) wird für Klonierungsexperimente angestrebt. Dabei wird der Hydrolysevorgang frühzeitig abgebrochen. Mehrfachverdaue, also die Verwendung von zwei oder mehreren REs werden für Fingerprintings und Schnittstellenkartierungen eingesetzt.
Manchmal wird bei Klonierungsexperimenten auf die Bildung so genannter kompatibler Enden geachtet. So bilden z.B. Aci I, Bsa H I, Cla I, Hap II, Mae II, Nar I und Tap I 5´CG-Überhänge in ihren Schnittstellen, die eine Klonierung erleichtern können.
Rekombinante Herstellung von Restriktionsenzymen
Viele Restriktionsendonukleasen werden heute nicht mehr aus ihrem ursprünglichen Produzenten isoliert, sondern sie werden in einen leicht handhabbaren Stamm kloniert.
Beispiel: Pst I aus Providencia stuartii in E. coli
Erst wird die Methylase (M.Pst I), dann die RE expremiert!
Verfahren:
- Chromosomale DNA aus P. stuartii mit Hind III schneiden
- Hind III-Fragment in Expressionsvektor ligieren
- Vektor in E.coli HB101 transformieren und im Flüssigmedium mit Lambda-Phagen infizieren (wenn RE rekombinant hergestellt wird, dann wird Lambda-DNA durch Pst I-produzierende und damit Lambda-resistente E. coli abgebaut)
- Osmotischer Schock Lambda-resistenter E. coli Zellen: Freisetzen der periplasmatischen Enzyme (inklusive des Pst I)
- Test auf Pst I
- Ergebnis: 10-fache Überexpression der Pst I gegenüber dem ursprünglichen Produzenten.